
Gliederung
2.1 Das Prinzip der chemischen Evolution
2.2 Theorieansätze
2.2.1 Oparin-Haldane-Hypothese – Die Entstehung des Lebens auf der Erde
2.2.2 Die Panspermie-Hypothese – Das Leben aus dem Weltall
3 Das Leben an extremen Standorten
3.1.1 Limitierende Faktoren in der Tiefsee
3.1.2 Anpassungsmechanismen von Lebewesen an die Tiefsee
3.2.1 Limitierende Faktoren im Hochgebirge
3.2.2 Anpassungsmechanismen von Lebewesen im Hochgebirge
5 Literatur- und Quellenverzeichnis
1 Einleitende Worte
Die Erde beherbergt eine Vielfalt an unterschiedlichen Lebewesen, die zum Teil extremen Bedingungen trotzen. Wie sehen diese Bedingungen am tiefsten und am höchsten Punkt der Erdoberfläche aus und wie ist das Überleben dort möglich? Doch zunächst stellt sich die Frage, wie und wo das Leben entstand.
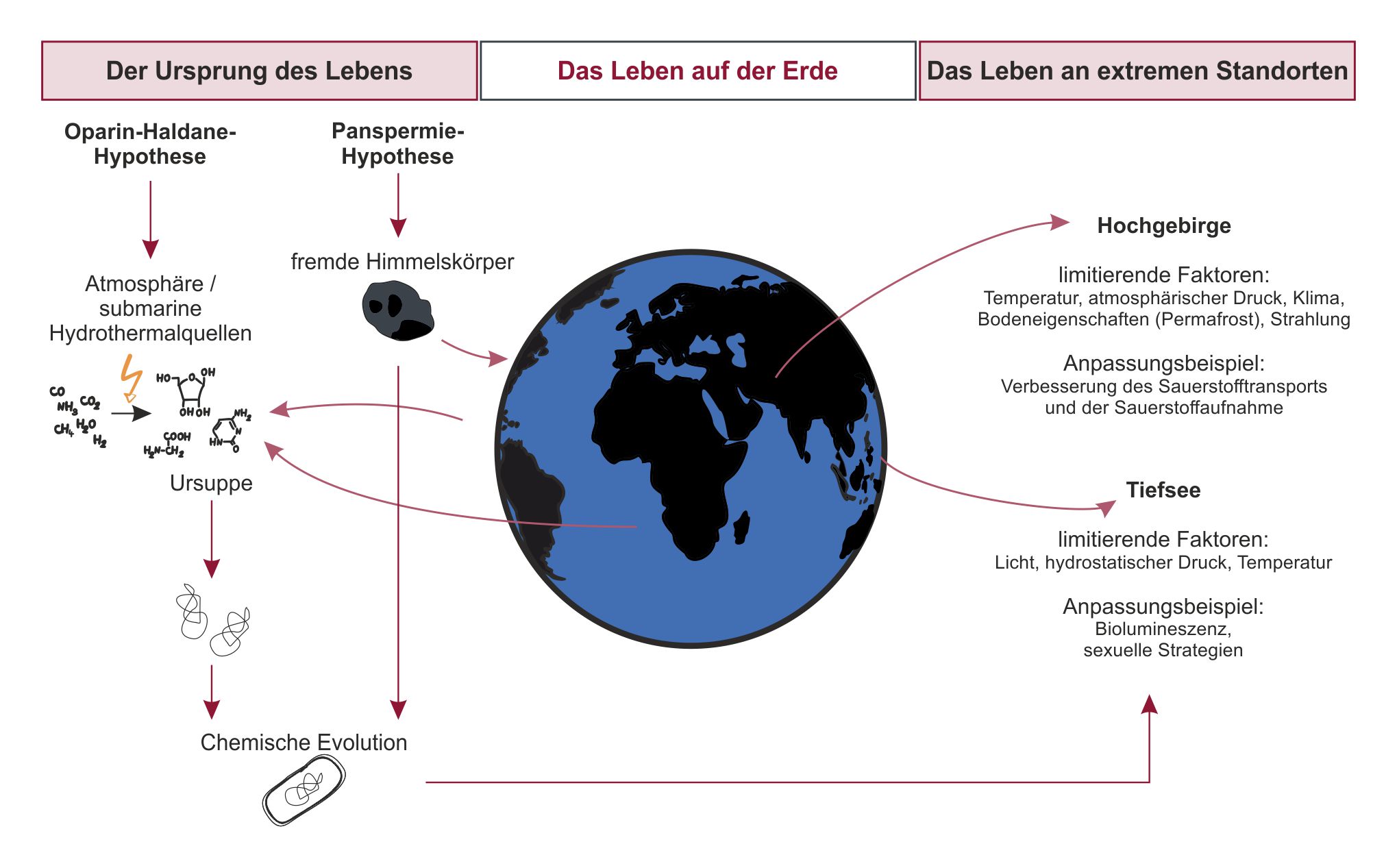
2 Der Ursprung des Lebens
Damit Leben entstehen konnte, muss einst aus anorganischem Material organisches entstanden sein, welches sich anschließend zu lebendigen Zellen entwickelte. Dieser Prozess wird als chemische Evolution bezeichnet (Calvin 1956: 387). Doch gibt es unterschiedliche Theorien zum genauen Ablauf und Ort.
2.1 Das Prinzip der chemischen Evolution
Erste Überlegungen zu der Entstehung des Lebens machte bereits Charles Darwin, Begründer der modernen Evolutionstheorie, die er 1871 in einem Brief äußerte (Darwin Correspondence Project). 50 Jahre später wurden konkrete Hypothesen über den Ursprung des Lebens formuliert und in den 1950er Jahren erste Experimente zu dem Thema entwickelt (Animation 1). Mit der daraus hervorgehenden Theorie der chemischen Evolution versuchte man die Entstehung von Grundbausteinen des Lebens aus nichtlebenden Systemen zu erklären. Aus chemischen Reaktionen gingen demnach biologische Strukturen wie Zucker, Nukleinbasen und Aminosäuren hervor, die die Grundbausteine von Zellen darstellen (Eschenmoser 2009: 182). Aus diesen Verbindungen konnten daraufhin durch weitere Reaktionen frühe Formen des Lebens entstehen (La Escosura 2019: 1).
2.2 Theorieansätze
Die Theorien zu der Entstehung des Lebens sind äußerst komplex und werden auch zur heutigen Zeit noch auf Grundlage neuer Erkenntnisse in den Naturwissenschaften kontinuierlich ergänzt und überholt (Kitadai & Maruyama 2018: 1144). Dennoch gibt es zwei grundlegende Ansätze zu unterscheiden.
2.2.1 Oparin-Haldane-Hypothese – Die Entstehung des Lebens auf der Erde
Eine gängige Theorie dazu wie das Leben entstanden sein könnte wurde in den 1920ern von zwei unterschiedlichen Wissenschaftlern unabhängig voneinander aufgestellt (Ponnamperuma 1964: 339). Diese sogenannte Oparin-Haldane-Hypothese besagt, dass alle Voraussetzungen für die Entstehung des Lebens auf der Urerde gegeben waren (Fry 2006: 25). Die chemische Evolution wurde demnach durch die Abkühlung der Erde nach ihrer Differenzierung vor etwa 3,8 Milliarden Jahren initiiert. Die Abkühlung hatte die Entstehung der Erdkruste sowie die Kondensation von Wasser zur Folge, wodurch sich Ozeane bildeten (Tirard 2017: 736 aus Haldane 1929). Zudem hatte die Atmosphäre zur damaligen Zeit eine andere Zusammensetzung als heute. Unter anderem bestand sie aus Ammoniak und großen Mengen an Kohlenstoffdioxid, enthielt jedoch keinen Sauerstoff (Kasting 1993: 921). Die Zusammensetzung dieser jungen Atmosphäre soll im ersten Schritt der Entstehung des Lebens bewirkt haben, dass mehr chemisch aktive UV-Strahlung hindurchdringen konnte, als es bei der heutigen Erdatmosphäre der Fall ist. Die Strahlung soll in Verbindung mit Wärme als chemischer Katalysator auf das atmosphärische Gemisch aus Wasser, Kohlenstoffdioxid und Ammoniak gewirkt haben, woraus erste organische Substanzen entstanden (Tirard 2017: 736). Diese neuen Verbindungen haben sich laut der Hypothese in einem zweiten Schritt in den Urozeanen angereichert, sodass die Viskosität des Wassers stark zunahm. Diese als Ursuppe bezeichnete Mixtur hatte aufgrund der großen Menge an verschiedenen chemischen Verbindungen äußerst reaktive Eigenschaften. Laut der Theorie führte die Reaktivität unter dem anhaltenden Einwirken der Sonnenradiation und der daraus resultierenden Wärme dazu, dass größere organische, selbstreproduktive Moleküle entstanden. Sie schlossen sich dann vermehrt zufällig in öligen Filmen und Wasser zusammen und bildeten so im dritten Schritt primitive Urzellen (Animation 2) (Tirard 2017: 736).
Diese zunächst theoretische Hypothese wurde erstmalig im Jahr 1953 durch das sogenannte Miller-Urey-Experiment überprüft (Fry 2006: 24). Das Experiment bestand aus einer Apparatur, die den Zustand der Erde zur damaligen Zeit simulierte und so die in der Oparin-Haldane-Hypothese beschriebenen Reaktionen überprüfte (Urey 1952; Miller 1953; Video 1). Mit diesem und weiteren Experimenten konnte nachgewiesen werden, dass diese Hypothese eine Möglichkeit darstellt, wie der Ursprung des Lebens ausgesehen haben könnte (Fry 2006: 24).
Mit dem Fortschreiten der Wissenschaft wurde diese Theorie seitdem weiter ausgebaut. So ist beispielsweise bekannt, dass der Vulkanismus nicht nur einen wesentlichen Anteil an der Zusammensetzung der Atmosphäre zu Zeiten der chemischen Evolution hatte, sondern auch durch die Bildung von submarinen Hydrothermalquellen einen weiteren möglichen Ursprungsort des Lebens darstellt (Corliss et al. 1981: 60). Durch die hohen Temperaturen und die Zusammensetzung herausströmender Gase sind auch dort laut der Hypothese Bedingungen gegeben, aus denen organischen Verbindungen entstanden sein könnten (Miller & Bada 1988: 610).
2.2.2 Die Panspermie-Hypothese – Das Leben aus dem Weltall
Parallel zu der Theorie, dass der Lebensursprung auf der Erde selbst liegt, existiert die sogenannte Panspermie-Hypothese (Audio 1), die besagt, dass das Leben oder Vorläufer des Lebens in Form von Molekülen aus dem Weltall auf die Erde kamen (Bezverkhniy & Bezverkhniy 2020: 2). Auch hierzu gibt es unterschiedliche Überlegungen. Frühe Vertreter der Panspermie-Hypothese vermuteten, dass diese Lebensformen durch elektromagnetische Kräfte auf die Erde kamen (Arrhenius 1908) oder durch extraterrestrische intelligente Lebensformen aktiv auf die Erde gebracht wurden, um diese zu kolonialisieren (Crick & Orgel 1973: 342). Diese Überlegungen bildeten das Fundament für die vielfältigen Denkrichtungen der Panspermie-Hypothese. Im Grunde können viele Einteilungen der zahlreichen Theorien vorgenommen werden, beispielsweise dahingehend, dass organische Moleküle und Verbindungen als Vorläufer des Lebens auf die Erde kamen und dort aufgrund der bereits in Abschnitt 2.2.1 genannten Bedingungen zu lebenden Zellen wurden (Matthews & Minard 2006: 398) oder dass bereits lebendige Zellen möglicherweise in Form von Bakterien, Pilzen oder Viren durch Kollisionen mit anderen Himmelskörpern auf die Erde kamen (Hoyle & Wickramasinghe 1981: 230). Letztere Denkweise wird derzeit auch auf das Erscheinen des Coronavirus und anderer Krankheitserreger angewandt, da ein räumlicher und zeitlicher Zusammenhang zwischen einem Meteoriteneinschlag und dem Ausbruch des Virus entdeckt wurde (Steele et al. 2020: 88ff). Da auf ebensolchen Himmelkörpern bereits im Weltall organische Verbindungen nachgewiesen werden konnten (Wright et al. 2015) und gezeigt wurde, dass Bakterien über längere Zeiträume unter den dort gegebenen Bedingungen überlebensfähig sind (La Torre et al. 2010), ist auch diese Theorie eine ernst zu nehmende Möglichkeit, wie das Leben eventuell auf die Erde gekommen ist. So wurde 2020 ein Experiment veröffentlicht, bei dem ein Bakterium außerhalb der International Space Station (ISS) den Bedingungen des interplanetaren Raums ausgesetzt wurde und nach einer Prognose mehrere Jahrzehnte lang dort überleben könnte (Kawaguchi et al. 2020: 9).
3 Das Leben an extremen Standorten
Dass die Mikroorganismen der Erde in der Lage sind teils extremen Bedingungen standzuhalten, macht die Forschung zur Panspermie-Hypothese deutlich. Doch auch höhere Lebewesen haben sich an die Bedingungen der unterschiedlichen Lebensräume auf der Erde angepasst. Wie extrem diese Bedingungen teilweise sind und wie die jeweiligen Anpassungen daran aussehen können, soll anhand von zwei Beispielen verdeutlicht werden: Der Tiefsee und dem Hochgebirge.
3.1 Das Leben in der Tiefsee
3.1.1 Limitierende Faktoren in der Tiefsee
Die Tiefsee ist vor allem eins – dunkel. Denn durch die optischen Eigenschaften des Wassers wird bereits in geringen Tiefen der Großteil der solaren Strahlung absorbiert und gestreut. UV-Licht und Licht im Infrarot- und Rotbereich erreichen im offenen Ozean nur sehr geringe Tiefen von 10 m bis maximal 50 m während in Tiefen von über 100 m nur Blauanteile des Lichts mit Wellenlängen um 450 nm gelangen (Darwiesh et al. 2018: 3; Woźniak & Dera 2007: 3). Ab etwa 200 m unter der Wasseroberfläche ist fast kein Licht mehr vorhanden (Animation 3) (Prazeres & Renema 2019: 8). Der tiefste Punkt der Ozeane ist das Challengertief des Marianengrabens, das laut einer Messung im Jahr 2014 10.984 m von der Wasseroberfläche entfernt liegt (Gardner et al. 2014: 11) und tektonischen Ursprungs ist (Lemenkova 2018: 41). Somit gibt es einen großen Bereich, in dem Dunkelheit herrscht. Dieser Bereich wird als Bathyal oder midnight zone bezeichnet (van den Hoff et al. 2017: 1). Neben der Dunkelheit sind Lebewesen hier zudem einem extrem hohen hydrostatischen Druck und kalten Wassertemperaturen ausgesetzt (van den Hoff et al. 2017: 1). Das fehlende Licht hat neben dem visuellen Aspekt einen weiteren Einfluss. Denn nur mit Licht kann Photosynthese betrieben werden (Ryther 1956: 69), beispielsweise durch Algen, welche die Hauptproduzenten des im Ozean vorhandenen Sauerstoffs darstellen (Riser & Johnson 2008: 324). Somit steht im Bathyal neben den anderen limitierenden Faktoren auch weniger gelöster Sauerstoff zur Verfügung.
3.1.2 Anpassungsmechanismen von Lebewesen an die Tiefsee
Um die Komplexität der Anpassungsmechanismen von Lebewesen an diesen Lebensraum zu veranschaulichen, dient die Anpassung an den Faktor Licht als gutes Exempel. Wo kein Licht ist, ist auch die Suche nach Beute oder Fortpflanzungspartnern und somit die Reproduktion sowie Kommunikation der hier lebenden Arten erschwert (Davis et al. 2014: 1139). Aus diesem Grund sind viele der dort lebenden Arten dazu in der Lage über eine chemische Reaktion eines körpereigenen Enzyms selbst Licht zu produzieren (Widder 2010: 705, Video 2). Dieses Licht ist zumeist blau, da das visuelle System von Organismen in großen Tiefen an das noch weit nach unten reichende, blaue Lichtspektrum angepasst ist (Widder 2010: 705 aus Archer et al. 1999). Nicht nur die Rezeption von Farben ist an die Bedingungen der Tiefsee angepasst. Viele dort lebende Tiere haben auch besonders große oder sensitive Augen ausgebildet, um möglichst viel Restlicht einfangen zu können (Warrant & Locket 2004: 677; Nilsson et al. 2012: 683). Trotz der Befähigung zur Biolumineszenz treffen sich seltene Spezies nicht häufig untereinander. Daher haben sich teilweise besondere sexuelle Strategien entwickelt, so wie der sexuelle Parasitismus des Tiefsee-Anglerfischs (Caulophryne jordani) (Pietsch 2005: 207). Das Männchen dieser Art ist deutlich kleiner als das Weibchen. Finden sich zwei Individuen der beiden Geschlechter, heftet sich das Männchen mit seinem Maul an das Weibchen und die Gewebe beider Anglerfische verbinden sich. So wird das Männchen lebenslang über den Blutkreislauf des Weibchens mit lebensnotwendigen Nährstoffen versorgt und das Weibchen sichert sich eine Samenquelle (Pietsch 2005: 207). Forschern der Rebikoff Foundation gelang es 2018 erstmalig, ein solches Paar in 800 m Tiefe zu filmen (News at a glance 2018: 1311, Video 3).
3.2 Das Leben im Hochgebirge
3.2.1 Limitierende Faktoren im Hochgebirge
Ein markantes Merkmal der Gebirge ist die abnehmende atmosphärische Temperatur mit zunehmender Höhe (Abbildung 2), durch die es auch zu Wechseln des Klimas kommt. Das Klima beeinflusst wiederum maßgeblich die Vegetation (Derguy et al. 2019: 3). Um Biomasse aufbauen zu können, benötigen Pflanzen je nach Art gewisse Minimaltemperaturen in ihrer Vegetationsperiode aber auch chemisch-physikalische Bodeneigenschaften, die ein Wachstum ermöglichen (Polis 1999: 4). Die Bedingungen spiegeln sich im Höhenprofil von Gebirgen wider, wo die Vegetation oberhalb der Wald- und der Baumgrenze mit Einsetzen der Schneegrenze nicht mehr vorhanden ist (Rubel et al. 2017: 121). Weitere Faktoren, die in Hochgebirgen die Vegetation beeinflussen sind das Mitreißen von Pflanzen durch Lawinen, die schlechten Bodeneigenschaften durch Permafrost, die erhöhte Strahlung aufgrund der dünneren schützenden Atmosphäre und der Reflektion von Strahlung durch die Schneedecke sowie erhöhte Windgeschwindigkeiten (Bebi et al. 2004: 208). In welcher Höhe diese Bedingungen eintreten hängt von der geographischen Lage der Gebirge und dem dort jeweils herrschenden Klima ab (Derguy et al. 2019: 3; Animation 4).
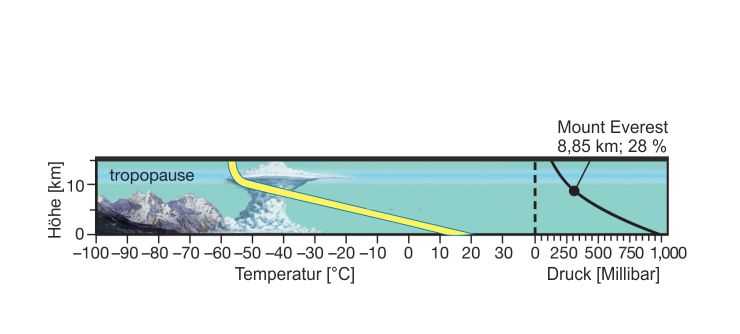
Ein anderes Merkmal, das wiederum die Tierwelt sowie den Menschen stark beeinflusst, ist neben den niedrigen Temperaturen in hohen Lagen der abnehmende Luftdruck (Burrows et al. 2011: 9). Auf dem mit 8.848 m höchsten Berg der Erde, dem Mount Everest, beträgt der atmosphärische Druck etwa ein Drittel des atmosphärischen Drucks auf Meeresniveau (West 1999: 1064). Durch das Einatmen von Luft geringen Drucks ist auch die Sauerstoffaufnahme eingeschränkt (Schoene 2001: 3121). In der sogenannten Todeszone ab 7.500 m Höhe ist der menschliche Körper durch den akuten Sauerstoffmangel so geschwächt, dass er hier nur wenige Stunden lebensfähig ist. Doch bereits ab einer Höhe von 2.500 m und insbesondere ab 5.500 m, wo der atmosphärische Druck nur 50 % des Drucks auf Meeresniveau entspricht, kann es zu Symptomen der Höhenkrankheit kommen. Hierzu zählen unter anderem Wassereinlagerungen in den Hirn- oder Lungengefäßen (Ödeme), die unbehandelt schnell zum Tod führen können. Durch eine Akklimatisierung des Körpers kann das Risiko jedoch stark minimiert werden und nur so ist das Erreichen großer Höhen überhaupt möglich (Pynn 2013: 141).
3.2.2 Anpassungsmechanismen von Lebewesen im Hochgebirge
Um den extremen Bedingungen der Hochgebirge besser standhalten zu können, haben einige Pflanzen besondere Merkmale ausgebildet. So gibt es eine Vielfalt von Arten, die sich an kalte Temperaturen, Trockenstress durch gefrorene Böden oder anderen Faktoren angepasst haben (Billings 1974: 132ff). Bei Tieren kann ein verbesserter Sauerstofftransport in Blut und Geweben festgestellt werden (Monge & Leon-Velarde 1991: 1137), was auch beim Menschen durch eine Höhenanpassung erreicht wird. Die Streifengans (Anser indicus) ist beispielsweise dazu in der Lage im Jetstream mit einer Geschwindigkeit von über 150 km/h das Himalaya-Gebirge zu überqueren (Newton 2008).
In den letzten Jahren wird zunehmend das Volk der Sherpa untersucht, welches Anzeichen physiologischer Anpassungen an die Höhe zeigt (Bhandari & Cavalleri 2019: 2ff), die wahrscheinlich durch eine Genmutation zustande kommen (Gnecchi-Ruscone et al. 2018: 2927; Hanaoka et al. 2012: 2).
4 Abschließende Worte
Das Leben ist so vielschichtig wie die physisch-geographischen Gegebenheiten der Erde sowie die globalen Unterschiede, an die es auf verschiedene Weisen angepasst ist. Die Komplexität wird bereits durch die Vielfalt der unterschiedlichen Entstehungstheorien des Lebens deutlich. Viele Wissenschaftler sind der Meinung, dass das Geheimnis um die Entstehung des Lebens vermutlich niemals gelüftet werden wird (Krishnamurthy & Hud 2020: 4614). Die zwei beschriebenen Theorieansätze sind jedoch anerkannte Möglichkeiten, wie es passiert sein könnte. Auch wenn die Hypothesen schon lange existieren, werden sie auch heute noch fortwährend aktualisiert.
5 Literatur- und Quellenverzeichnis
Archer, S.; Djamgoz, M. B.; Loew, E.; Partridge, J. C.; Vallerga, S. (1999): Adaptive Mechanisms in the Ecology of Vision. 1st ed. 1999. Dordrecht: Springer Netherlands : Imprint: Springer.
Arrhenius, Svante (1908): Worlds in the making. the evolution of the universe. New York, London: Harper & Brothers publishers. Online verfügbar unter https://sealevel.info/Arrhenius1908_worldsinmaking00.pdf, zuletzt geprüft am 04.01.2021.
Bebi, Peter; Casteller, Alejandro; Mayer, Andrea Corinna; Stöckli, Veronika (2004): Jahrringe als Indikatoren für extreme Standortsbedingungen im Gebirge: Schnee, Lawinen und Permafrost | Growth rings as indicators of extreme site conditions in the mountains: snow, avalanches and permafrost. In: Schweizerische Zeitschrift fur Forstwesen 155 (6), S. 208–212. DOI: 10.3188/szf.2004.0208  .
.
Bezverkhniy, Volodymyr; Bezverkhniy, Vitaliy (2020): The Origin of Life on Earth, the Panspermia Hypothesis and Cosmological DNA Synthesis. In: SSRN Electronic Journal. DOI: 10.2139/ssrn.3678305 .
Bhandari, Sushil; Cavalleri, Gianpiero L. (2019): Population History and Altitude-Related Adaptation in the Sherpa. In: Front. Physiol. 10, S. 1116. DOI: 10.3389/fphys.2019.01116 .
Billings, W. D. (1974): Adaptations and Origins of Alpine Plants. In: Arctic and Alpine Research 6 (2), S. 129–142. DOI: 10.1080/00040851.1974.12003769 .
Burrows, J. P.; Goede, A. P. H.; Muller, C.; Bovensmann, H. (2011b): SCIAMACHY – The Need for Atmospheric Research from Space. In: Manfred Gottwald und Heinrich Bovensmann (Hg.): SCIAMACHY – Exploring the Changing Earth’s Atmosphere. Dordrecht: Springer Science+Business Media B.V, S. 1–17. DOI: 10.1007/978-90-481-9896-2_1 ' fill='%235dd95d'%3E%3Cpath d='M 9.249787330627441 17.17169380187988 L 4.762476921081543 12.68437480926514 L 7.214956283569336 10.23189544677734 L 9.015677452087402 12.03262424468994 L 9.369227409362793 12.38618469238281 L 9.722777366638184 12.03262424468994 L 16.32264709472656 5.432744979858398 L 18.65010833740234 7.760213375091553 L 9.249787330627441 17.17169380187988 Z' stroke='none'/%3E%3Cpath d='M 9.249578475952148 16.46436309814453 L 17.94319725036621 7.760422229766846 L 16.32264709472656 6.139865875244141 L 10.07633686065674 12.38618469238281 L 9.369227409362793 13.09329414367676 L 8.662117004394531 12.38618469238281 L 7.214955806732178 10.93901634216309 L 5.469596385955811 12.68437480926514 L 9.249578475952148 16.46436309814453 M 9.249997138977051 17.87900352478027 L 4.05537748336792 12.68437385559082 L 7.214957237243652 9.524794578552246 L 9.369227409362793 11.67907428741455 L 16.32264709472656 4.725644111633301 L 19.35699653625488 7.76000452041626 L 9.249997138977051 17.87900352478027 Z' stroke='none' fill='%23fff'/%3E%3C/g%3E%3C/svg%3E%0A "In Citavi anzeigen") .
.
Calvin, Melvin (1956): Die chemische Evolution und der Ursprung des Lebens. In: Naturwissenschaften (17), S. 387–393.
Corliss, J. B.; Baross, Ja; Hoffman, Se (1981): An Hypothesis Concerning the Relationships Between Submarine Hot Springs and the Origin of Life on Earth. In: Oceanologica Acta, Special issue. Online verfügbar unter https://archimer.ifremer.fr/doc/00245/35661/, zuletzt geprüft am 02.01.2021.
Crick, Francis H. C.; Orgel, Leslie E. (1973): Directed panspermia. In: Icarus 19 (3), S. 341–346. DOI: 10.1016/0019-1035(73)90110-3 .
Darwiesh, M.; El-Sherif, Ashraf; Ayoub, H.; El-sharkawy, Yasser; Hassan, Mahmoud (2018): The Design and Implementation of Under-Water Laser Imaging Test Aquarium. In: The International Conference on Mathematics and Engineering Physics 9 (6), S. 1–15. DOI: 10.21608/icmep.2018.29582 .
Darwin Correspondence Project: Letter no. 7471. University of Cambridge. Cambridge. Online verfügbar unter https://www.darwinproject.ac.uk/letter/DCP-LETT-7471.xml, zuletzt geprüft am 11.12.2020.
Davis, Matthew P.; Holcroft, Nancy I.; Wiley, Edward O.; Sparks, John S.; Leo Smith, W. (2014): Species-specific bioluminescence facilitates speciation in the deep sea. In: Marine biology 161 (5), S. 1139–1148. DOI: 10.1007/s00227-014-2406-x .
Derguy, María R.; Frangi, Jorge L.; Drozd, Andrea A.; Arturi, Marcelo F. (2019): Holdridge Life Zone Map: Republic of Argentina. San Juan, Puerto Rico: International Institute of Tropical Forestry.
Encyclopedia Britannica (2012): Stratopause | meteorology. Online verfügbar unter https://www.britannica.com/science/stratopause, zuletzt geprüft am 09.01.2021.
Eschenmoser, Albert (2009): The search for the chemistry of life’s origin. In: Pontifical Academy of Sciences 20, S. 181–199. Online verfügbar unter http://www.pas.va/content/dam/accademia/pdf/acta20/acta20-eschenmoser.pdf, zuletzt geprüft am 04.01.2021.
Freistetter, Florian (2015): Sternengeschichten Folge 123: Panspermie – Kam das Leben aus dem Weltall – Florian Freistetter – Astrodicticum Simplex. Online verfügbar unter https://florian-freistetter.de/podcast/sternengeschichten-folge-123-panspermie-kam-das-leben-aus-dem-weltall/, zuletzt geprüft am 16.02.2021.
Fry, Iris (2006): The origins of research into the origins of life. In: Endeavour 30 (1), S. 24–28. DOI: 10.1016/j.endeavour.2005.12.002 .
Gardner, James V.; Armstrong, Andrew A.; Calder, Brian R.; Beaudoin, Jonathan (2014): So, How Deep Is the Mariana Trench? In: Marine Geodesy 37 (1), S. 1–13. DOI: 10.1080/01490419.2013.837849 .
Gnecchi-Ruscone, Guido A.; Abondio, Paolo; Fanti, Sara de; Sarno, Stefania; Sherpa, Mingma G.; Sherpa, Phurba T. et al. (2018): Evidence of Polygenic Adaptation to High Altitude from Tibetan and Sherpa Genomes. In: Genome Biol Evol 10 (11), S. 2919–2930. DOI: 10.1093/gbe/evy233 .
Haldane, John B. S. (1929): The Origin of life. In: The Rationalist Annual 148, S. 3–10.
Hanaoka, Masayuki; Droma, Yunden; Basnyat, Buddha; Ito, Michiko; Kobayashi, Nobumitsu; Katsuyama, Yoshihiko et al. (2012): Genetic variants in EPAS1 contribute to adaptation to high-altitude hypoxia in Sherpas. In: PloS one 7 (12), e50566. DOI: 10.1371/journal.pone.0050566 .
Hoyle, Fred; Wickramasinghe, Chandra (1981): Comets – A Vehicle for Panspermia. In: Cyril Ponnamperuma (Hg.): Comets and the Origin of Life. Proceedings of the Fifth College Park Colloquium on Chemical Evolution, University of Maryland, College Park, Maryland, U.S.A., October 29th to 31st, 1980. Dordrecht, 1981. Dordrecht: Springer (Proceedings of the College Park Colloquia, 5), S. 227–239.
Junker, Reinhard; Scherer, Siegfried (2006): Evolution: Ein kritisches Lehrbuch. 6. Aufl. Gießen: Weyel.
Kasting, James F. (1993): Earth’s early atmosphere. In: Science 259 (5097), S. 920–926. DOI: 10.1126/science.11536547 .
Kawaguchi, Yuko; Shibuya, Mio; Kinoshita, Iori; Yatabe, Jun; Narumi, Issay; Shibata, Hiromi et al. (2020): DNA Damage and Survival Time Course of Deinococcal Cell Pellets During 3 Years of Exposure to Outer Space. In: Front. Microbiol. 11. DOI: 10.3389/fmicb.2020.02050 .
Kitadai, Norio; Maruyama, Shigenori (2018): Origins of building blocks of life: A review. In: Geoscience Frontiers 9 (4), S. 1117–1153. DOI: 10.1016/j.gsf.2017.07.007 .
Krishnamurthy, Ramanarayanan; Hud, Nicholas V. (2020): Introduction: Chemical Evolution and the Origins of Life. In: Chemical reviews 120 (11), S. 4613–4615. DOI: 10.1021/acs.chemrev.0c00409 .
La Escosura, Andrés de (2019): The Informational Substrate of Chemical Evolution: Implications for Abiogenesis. In: Life (Basel, Switzerland) 9 (3). DOI: 10.3390/life9030066 .
La Torre, Rosa de; Sancho, Leopoldo G.; Horneck, Gerda; Ríos, Asunción de los; Wierzchos, Jacek; Olsson-Francis, Karen et al. (2010): Survival of lichens and bacteria exposed to outer space conditions – Results of the Lithopanspermia experiments. In: Icarus 208 (2), S. 735–748. DOI: 10.1016/j.icarus.2010.03.010 .
Lemenkova, Polina (2018): R scripting libraries for comparative analysis of the correlation methods to iden- tify factors affecting Mariana Trench formation. In: Journal of Marine technology and Environment 2, S. 35–42. DOI: 10.6084/m9.figshare.7434167.v1 .
Matthews, Clifford N.; Minard, Robert D. (2006): Hydrogen cyanide polymers, comets and the origin of life. In: Faraday Discussions 133 (0), S. 393–401. DOI: 10.1039/B516791D .
Miller, Stanley L. (1953): A production of amino acids under possible primitive earth conditions. In: Science (New York, N.Y.) 117 (3046), S. 528–529. DOI: 10.1126/science.117.3046.528 .
Miller, Stanley L.; Bada, Jeffrey L. (1988): Submarine hot springs and the origin of life. In: Nature 334 (18), S. 609–611. Online verfügbar unter https://www.nature.com/articles/334609a0.pdf?origin=ppub, zuletzt geprüft am 02.01.2021.
Monge, Carlos; Leon-Velarde, Fabiola (1991): Physiological adaptation to high altitude: oxygen transport in mammals and birds. In: Physiological Reviews 71 (4), S. 1135–1172. DOI: 10.1152/physrev.1991.71.4.1135 .
News at a glance (2018). In: Science 359 (6382), S. 1310–1312. Online verfügbar unter https://science.sciencemag.org/content/359/6382/1310/tab-pdf.
Newton, Ian (2008): Chapter 6 – Incredible journeys. In: Ian Newton und Keith Brockie (Hg.): The migration ecology of birds. Amsterdam, London: Elsevier-Academic Press, S. 139–161. Online verfügbar unter https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/bar-headed-goose, zuletzt geprüft am 07.01.2021.
Nilsson, Dan-Eric; Warrant, Eric J.; Johnsen, Sönke; Hanlon, Roger; Shashar, Nadav (2012): A Unique Advantage for Giant Eyes in Giant Squid. In: Current Biology 22 (8), S. 683–688. DOI: 10.1016/j.cub.2012.02.031 .
Pietsch, Theodore W. (2005): Dimorphism, parasitism, and sex revisited: modes of reproduction among deep-sea ceratioid anglerfishes (Teleostei: Lophiiformes). In: Ichthyol Res 52 (3), S. 207–236. DOI: 10.1007/s10228-005-0286-2 .
Polis, Gary A. (1999): Why Are Parts of the World Green? Multiple Factors Control Productivity and the Distribution of Biomass. In: Oikos 86 (1), S. 3–15. DOI: 10.2307/3546565 .
Ponnamperuma, Cyril (1964): Chemical Evolution and the Origin of Life. In: Nature 201 (4917), S. 337–340. DOI: 10.1038/201337a0 .
Prazeres, Martina; Renema, Willem (2019): Evolutionary significance of the microbial assemblages of large benthic Foraminifera. In: Biological reviews of the Cambridge Philosophical Society 94 (3), S. 828–848. DOI: 10.1111/brv.12482 .
Pynn, Harvey (2013): ABC of Prehospital Emergency Medicine: BMJ Books. Online verfügbar unter https://books.google.de/books?id=Aa0qAAAAQBAJ&printsec=frontcover&hl=de#v=onepage&q&f=false, zuletzt geprüft am 07.01.2021.
Riser, Stephen C.; Johnson, Kenneth S. (2008): Net production of oxygen in the subtropical ocean. In: Nature 451 (7176), S. 323–325. DOI: 10.1038/nature06441 .
Rubel, Franz; Brugger, Katharina; Haslinger, Klaus; Auer, Ingeborg (2017): The climate of the European Alps: Shift of very high resolution Köppen-Geiger climate zones 1800–2100. In: metz 26 (2), S. 115–125. DOI: 10.1127/metz/2016/0816 .
Ryther, John H. (1956): Photosynthesis in the Ocean as a Function of Light Intensity. In: Limnology and Oceanography 1, S. 61–70.
Schoene, Robert B. (2001): Limits of human lung function at high altitude. In: The Journal of experimental biology 204 (Pt 18). Online verfügbar unter https://pubmed.ncbi.nlm.nih.gov/11581325/.
Science | AAAS (2018): Exclusive: ‘I’ve never seen anything like it.’ Video of mating deep-sea anglerfish stuns biologists. Online verfügbar unter https://www.sciencemag.org/news/2018/03/exclusive-i-ve-never-seen-anything-it-video-mating-deep-sea-anglerfish-stuns-biologists, zuletzt geprüft am 16.02.2021.
Steele, Edward J.; Gorczynski, Reginald M.; Lindley, Robyn A.; Tokoro, Gensuke; Temple, Robert; Wickramasinghe, N. Chandra (2020): Origin of new emergent Coronavirus and Candida fungal diseases-Terrestrial or cosmic? In: Advances in genetics 106, S. 75–100. DOI: 10.1016/bs.adgen.2020.04.002 .
Tirard, Stéphane (2017): J. B. S. Haldane and the origin of life. In: Journal of genetics 96 (5), S. 735–739. DOI: 10.1007/s12041-017-0831-6 .
Urey, Harold C. (1952): On the Early Chemical History of the Earth and the Origin of Life. In: Proceedings of the National Academy of Sciences of the United States of America 38 (4), S. 351–363. DOI: 10.1073/pnas.38.4.351 .
UW News (2018): A blind date in the deep sea: First-ever observations of a living anglerfish, a female with her tiny mate, coupled for life. Online verfügbar unter https://www.washington.edu/news/2018/03/22/a-blind-date-in-the-deep-sea-first-ever-observations-of-a-living-anglerfish-a-female-with-her-tiny-mate-coupled-for-life/, zuletzt geprüft am 18.01.2021.
van den Hoff, John; Kilpatrick, Robbie; Welsford, Dirk (2017): Southern elephant seals (Mirounga leonina Linn.) depredate toothfish longlines in the midnight zone. In: PloS one 12 (2), e0172396. DOI: 10.1371/journal.pone.0172396 .
Warrant, Eric J.; Locket, N. Adam (2004): Vision in the deep sea. In: Biological Reviews 79 (3), S. 671–712. DOI: 10.1017/S1464793103006420 .
West, John B. (1999): Barometric pressures on Mt. Everest: new data and physiological significance. In: Journal of applied physiology (Bethesda, Md. : 1985) 86 (3), S. 1062–1066. DOI: 10.1152/jappl.1999.86.3.1062 .
Widder, Edith A. (2010): Bioluminescence in the Ocean: Origins of Biological, Chemical, and Ecological Diversity. In: Science 328 (5979), S. 704–708. DOI: 10.1126/science.1174269 .
Woźniak, Bogdan; Dera, Jerzy (2007): Light absorption in sea water. New York NY: Springer (Atmospheric and oceanographic sciences library, 33).
Wright, I. P.; Sheridan, S.; Barber, S. J.; Morgan, G. H.; Andrews, D. J.; Morse, A. D. (2015): CHO-bearing organic compounds at the surface of 67P/Churyumov-Gerasimenko revealed by Ptolemy. In: Science 349 (6247). DOI: 10.1126/science.aab0673 .